
| 1) Optimization of goal-directed movements: |
|---|
|
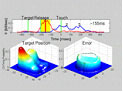 Voluntary goal-directed movements, such as arm reaching, are nearly optimized in terms of smoothness over the entire movement. Such smoothness is lost with cerebellar dysfunction, suggesting the essential role of the cerebellum in optimizing movement. However, it is still not clear how the cerebellum contributes to achieving smoothness over an entire movement. We have recently proposed a random walk hypothesis (Kitazawa 2002) that the terminal errors conveyed by climbing fibers in the cerebellum (Kitazawa et al. 1998) serve to reduce not only the mean error, but also the variance of the error, through a process analogous to the random walk through movement control candidates. We are planning electrophysiological experiments for testing this hypothesis.
Voluntary goal-directed movements, such as arm reaching, are nearly optimized in terms of smoothness over the entire movement. Such smoothness is lost with cerebellar dysfunction, suggesting the essential role of the cerebellum in optimizing movement. However, it is still not clear how the cerebellum contributes to achieving smoothness over an entire movement. We have recently proposed a random walk hypothesis (Kitazawa 2002) that the terminal errors conveyed by climbing fibers in the cerebellum (Kitazawa et al. 1998) serve to reduce not only the mean error, but also the variance of the error, through a process analogous to the random walk through movement control candidates. We are planning electrophysiological experiments for testing this hypothesis.
- Kitazawa, S, Wolpert, DM. Rhythmicity, randomness and synchrony in climbing fiber signals. Trends Neurosci. 2005;
28(11) 611-619.
- Kitazawa S. Optimization of goal-directed movements in the cerebellum: a random walk hypothesis. Neurosci Res 2002;43(4):289-94.
- Kitazawa S, Kimura T, Yin PB. Cerebellar complex spikes encode both destinations and errors in arm movements. Nature 1998;392(6675):494-7.
|
| 2) Neural representation of temporal order and simultaneity: |
|---|
|
 It is generally accepted that the brain can resolve the order of two stimuli that are separated in time by as little as 30 ms. This basically applies to temporal order judgments of two tactile stimuli, delivered one to each hand, as long as the arms are uncrossed. However, we recently found that crossing the arms caused many subjects to misreport (that is, invert) the temporal order (Yamamoto and Kitazawa 2001a). When the stimuli were delivered to the tips of sticks held in each hand, the judgment was dramatically altered by crossing the sticks without changing the positions of the hands (Yamamoto and Kitazawa 2001b). We suggest that it is not until the sensory signals are referred to relevant locations in space, which could be the hand itself or the tip of the stick in hand, that the stimuli are ordered in time. We are further examining the neural representation of temporal order with animals that are trained to judge temporal order of tactile stimuli.
It is generally accepted that the brain can resolve the order of two stimuli that are separated in time by as little as 30 ms. This basically applies to temporal order judgments of two tactile stimuli, delivered one to each hand, as long as the arms are uncrossed. However, we recently found that crossing the arms caused many subjects to misreport (that is, invert) the temporal order (Yamamoto and Kitazawa 2001a). When the stimuli were delivered to the tips of sticks held in each hand, the judgment was dramatically altered by crossing the sticks without changing the positions of the hands (Yamamoto and Kitazawa 2001b). We suggest that it is not until the sensory signals are referred to relevant locations in space, which could be the hand itself or the tip of the stick in hand, that the stimuli are ordered in time. We are further examining the neural representation of temporal order with animals that are trained to judge temporal order of tactile stimuli.
- Wada M, Moizumi S, Kitazawa S. Temporal order judgment in mice. Behav Brain Res 2005;157(1):167-175.
- Wada M, Yamamoto S, Kitazawa S. Effects of handedness on tactile temporal order judgment. Neuropsychologia 2004;42(14):1887-95.
- Kitazawa S. Where conscious sensation takes place. Conscious Cogn 2002;11(3):475-7.
- Yamamoto S, Kitazawa S. Reversal of subjective temporal order due to arm crossing. Nat Neurosci 2001;4(7):759-65.
- Yamamoto S, Kitazawa S. Sensation at the tips of invisible tools. Nat Neurosci 2001;4(10):979-80.
|
| 3) The role of dopamine and serotonin in the basal ganglia: |
|---|
|
 Monoaminerigic neurons (dopamine, serotonin) are known to be very important in learning, but their roles are not still characterized enough. Using in vivo voltammetry system which enables simultaneous measurement of dopamine and serotonin at 4 Hz for more than one year, the role of these transmitters in learning is studied. Also, we study the mechanism of Parkinson�fs disease from the neuropharmacological aspects, and study the function of trace amine (tyramine) in the brain.
Monoaminerigic neurons (dopamine, serotonin) are known to be very important in learning, but their roles are not still characterized enough. Using in vivo voltammetry system which enables simultaneous measurement of dopamine and serotonin at 4 Hz for more than one year, the role of these transmitters in learning is studied. Also, we study the mechanism of Parkinson�fs disease from the neuropharmacological aspects, and study the function of trace amine (tyramine) in the brain.
- Nakazato T, Akiyama A: Behavioral activity and stereotypy in rats induced by L-DOPA metabolites: a possible role in the adverse effects of chronic L-DOPA treatment of Parkinson's disease. Brain Res, 930: 134-142, 2002
- Nakazato T: The medial prefrontal cortex mediates 3-methoxytyramine-induced behavioural changes in rat. Eur J Pharmacol, 442: 73-79, 2002
- Nakazato T: Striatal dopamine release in the rat during a cued lever-press task for food reward and the development of changes over time measured using high-speed voltammetry. Exp. Brain Res, in press, 2005
|
| 4) Neural Mechanisms of Visual Perception: |
|---|
|
Understanding the neural mechanisms of visual perception is a major challenge in cognitive neuroscience. In this lab, we study the perception of motion and depth using psychophysical and neurophysiological methods. Our aim is to understand how visual information is encoded in the brain, and how they are read out to perform complex behavior.
- Uka T, DeAngelis GC. Contribution of area MT to depth discrimination: choice-related response modulations reflect task strategy. Neuron. 2004; 42: 297-310.
- Uka T, DeAngelis GC. Contribution of middle temporal area to coarse depth discrimination: comparison of neuronal and psychophysical sensitivity.J Neurosci. 2003; 23: 3515-3530.
- Uka T, Tanaka H, Yoshiyama K, Kato M, Fujita I. Disparity selectivity of neurons in monkey inferior temporal cortex. J Neurophysiol. 2000; 84: 120-132.
|
|

